エピジェネティック修飾塩基簡易測定法の開発
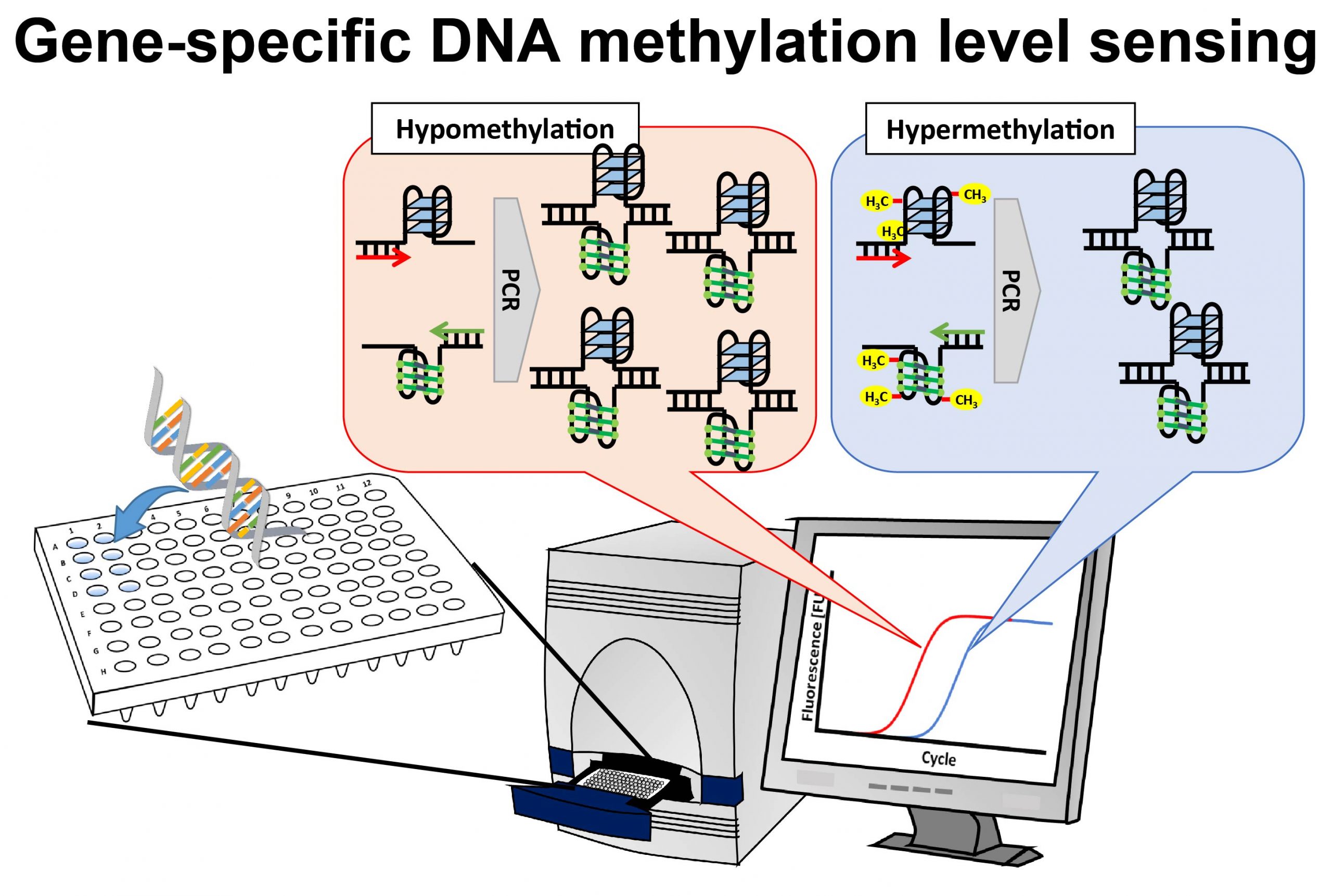
本研究室ではエピジェネティック修飾と呼ばれるDNAの修飾塩基に着目し、種々の疾患を簡便に診断できる技術の開発に取り組んでおります。エピジェネティック修飾塩基の中でメチルシトシンは最もよく解析されている修飾であり、遺伝子の発現を制御する遺伝子スイッチとして機能しております。がんや中枢神経疾患など疾患に関連する細胞では、この遺伝子スイッチ状態が異常になっているため、これを検査すれば種々の疾病の診断が可能になります。これまでに、メチル化されるとDNAの物理化学的性質が変化することに着目した遺伝子特異的メチル化レベル簡易測定法[37, 60]や、メチル化DNAに結合するヒトタンパク質とホタルの発光タンパク質を融合させた人工タンパク質を利用したゲノムDNA全体のメチル化レベル簡易測定法[38, 43, 48, 50, 51]、メチルシトシンの脱メチル化反応に関与しているTen-eleven translocation (TET) タンパク質を利用したゲノムDNA全体のメチル化レベル簡易測定法を開発しました[54]。さらに、脱メチル化反応の中間産物であるヒドロキシメチルシトシンを測定する方法も開発しました[59]。


DNA四重鎖構造の機能解析
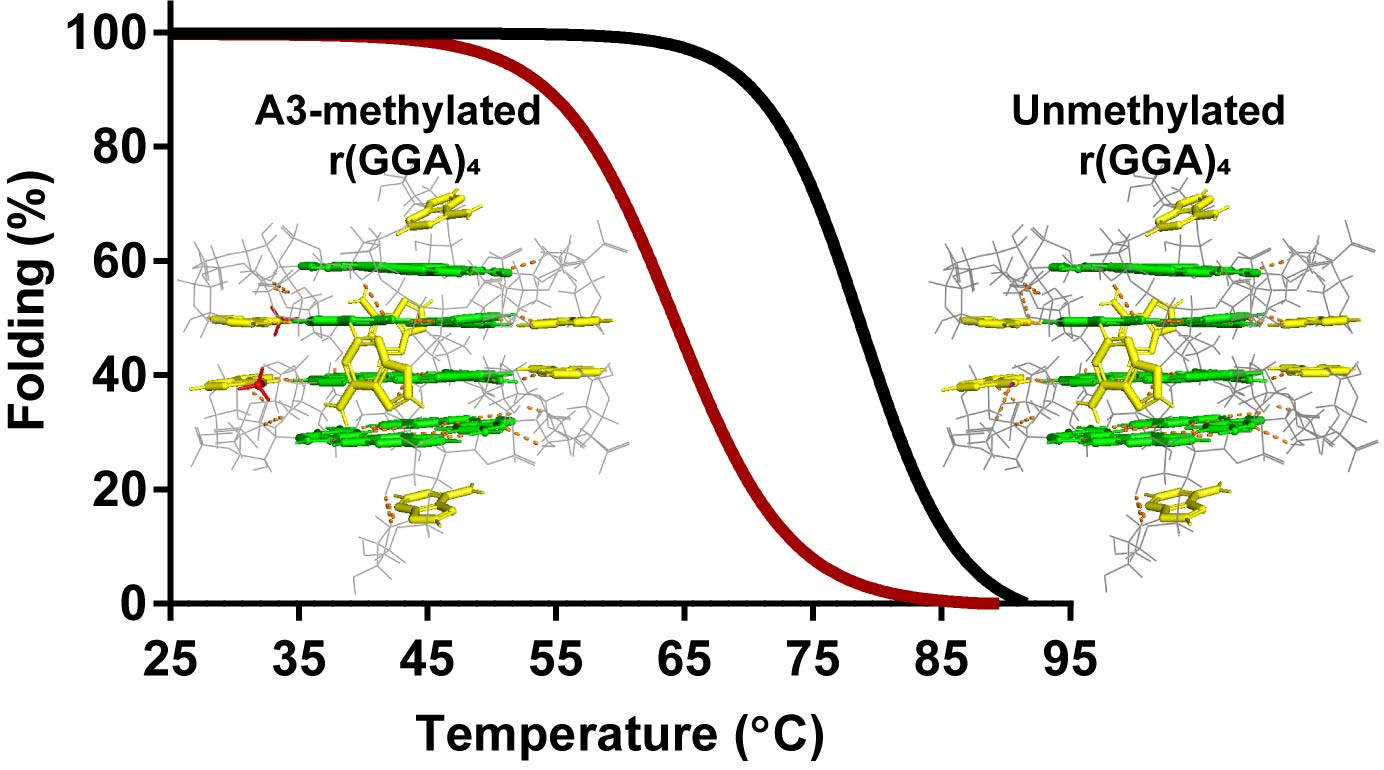
DNAは通常二重らせん構造を形成しますが、特定の配列を持つDNAはグアニン四重鎖(G-quadruplex: G4)構造やi-motif構造と呼ばれる特殊な高次構造を形成します。本研究室ではヒトゲノムDNA中でG4構造が形成される領域の解析やG4やi-motif構造形成が生命現象に与える影響を解析しております[42, 52]。これまでに次世代シークエンサーを利用してヒトゲノム中には9,651箇所のG4構造クラスターが存在することを明らかにしました[44]。また、エピジェネティック修飾がG4やi-motif構造形成に与える影響も解析しております。がん細胞などで過剰に発現している VEGF遺伝子やc-KIT遺伝子中に含まれるG4やimotif構造中のシトシンをメチル化すると、その熱安定性が変化することを明らかにしました[46, 56, 57]。さらに、c-KIT遺伝子やヒト染色体の末端領域であるテロメアで形成されるG4構造、GGA反復配列が形成するG4構造中のアデニンをメチル化すると、その熱安定性が変化することを明らかにしました[53, 55, 58]。